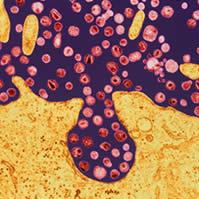
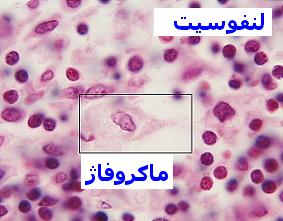
ماکروفاژ
مقدمه
اولین عملی که در ارتباط با سیستم ایمنی بدن به وقوع میپیوندند عبارت است از به داماندازی و نابود کردن و به عبارتی فاگوسیت کردن هر ماده خارجی که وارد بدن میشود. سلولهایی که مسئول این عمل در بدن پستانداران میباشند در دو سیستم طبقهبندی میشوند. اول سیستم میلویید که شامل سلولهایی است که دارای عمل سریع بوده ولی قادر به ادامه این عمل به مدت طولانی نیستند و شامل نوتروفیلها ، ائوزینوفیلها و بازوفیلها میباشد. دوم سلولهای سیستم فاگوسیتیک تکهستهای که کندتر عمل کرده ولی در عوض بارها عمل فاگوسیتوز را تکرار میکنند. این سلولها مسوول پردازش آنتیژن برای پاسخ ایمنی هستند. |
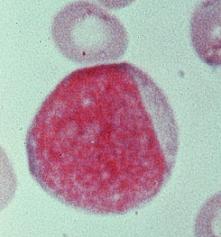
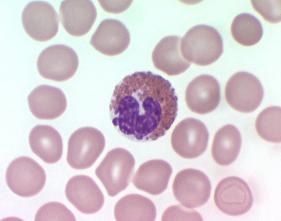
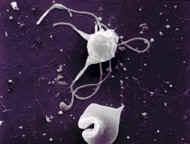
سیستم فاگوسیتیک تکهستهای از سلولهایی به نام ماکروفاژ که دارای یک هسته مدورند، تشکیل شده است. برخلاف نوتروفیلها ، ماکروفاژها قادر به انجام فاگوسیتوز بطور مداوم هستند. ماکروفاژها ، آنتیژن را مورد پردازش قرار داده و آنرا برای پاسخ ایمنی مهیا میسازند. این سلولها همچنین با از بین بردن بافتهای مرده ، در حال مرگ و یا تخریب شده مستقیما در عمل ترمیم نسوج شرکت مینمایند.
حضور ماکروفاژها در بافتهای مختلف بدن
ماکروفاژها انتشار وسیعی در سرتاسر بدن دارند. ماکروفاژهای نابالغ موجود در جریان خون به مونوسیت موسومند. مونوسیتها بطور معمول حدود 5% از جمعیت لوکوسیتها را تشکیل میدهند. ماکروفاژهای بالغ موجود در بافت پیوندی هیستوسیت (Histiocyte) نام دارد و آنهایی که جدار سینوزوییدهای کبد را میپوشانند به سلولهای کوپفر (Kupffer cells) موسومند.ماکروفاژهای موجود در مغز میکروگلیا (Microglia) و بالاخره آنهایی که در ریهها وجود دارند، ماکروفاژهای آلوئولار نامیده میشوند. تعداد زیادی از ماکروفاژها در طحال ، مغز استخوان و عقدههای لنفاوی ساکن هستند. صرفنظر از نام و مکان ، تمام این سلولها ماکروفاژ بوده و جزیی از سیستم فاگوسیتیک تکهستهای به شمار میروند.
ساختمان ماکروفاژها
شکل ظاهری ماکروفاژها با توجه به تنوع محل استقرار آنها در بدن متفاوت است. در سوسپانسیون ، آنها به شکل سلولهایی مدور با قطر تقریبی 14-20 میکرومتر هستند. ماکروفاژها دارای سیتوپلاسم فراوان و یک هسته واحد در مرکزشان میباشند. این هسته ممکن است گرد و یا لوبیایی شکل باشد. سیتوپلاسم اطراف هسته واجد میتوکندری ، تعداد زیادی لیزوزوم ، مقداری شبکه آندوپلاسمی دانهدار (خشن) و دستگاه گلژی میباشد و این نشاندهنده توانایی سلول برای سنتز و ترشح پروتئینهاست. سیتوپلاسم محیطی معمولا فاقد ارگانلهای سلولی بوده و دائما در حرکت است. به حرکت در آوردن این جز سیتوپلاسم با ایجاد مداوم پاهای کاذب به انجام میرسد.ماکروفاژها خیلی محکم به سطوح شیشهای چسبیده و سپس با خارج ساختن فیلامانهای سیتوپلاسمی باریک و طویل روی این سطوح پخش میشوند. بعضی ماکروفاژها دارای اختلافاتی با ساختمان پایه فوقالذکر میباشند. مثلا مونوسیتهای خون محیطی دارای هستههایی گرد هستند که با پیشرفت روند بلوغ سلول به تدریج از حالت گرد خارج شده و طویل میگردند، ماکروفاژهای آلوئولار معمولا فاقد شبکه آندوپلاسمیک دانهدار بوده و در عوض سیتوپلاسمشان پر از گرانول است و بالاخره سلولهای میکروگلیا موجود در سیستم عصبی مرکزی دارای هسته میلهای شکل و زواید سیتوپلاسمی طویلی هستند که این زواید در صورت تحریک سلول توسط آسیب بافتی از بین میروند.
 |
پاسخ ایمنی سلولی به برخی از ارگانیسمها سبب رشد ماکروفاژها و نیز افزایش تعداد لیزوزومهای موجود در آنها میگردد. سلولهایی که بدین طریق حاصل میشوند، ماکروفاژ فعال شده نام دارند. در صورتی که ماده بیگانه برای مدت طولانی در بدن باقی بماند، تعداد زیادی ماکروفاژ در اطراف این ماده تجمع حاصل کرده و از نظر بافتشناسی منظرهای شبیه به این تلیوم پیدا میکنند. از اینرو ، این سلولها به سلولهای اپیتلیویید موسومند. در صورتی که لزوم به دربرگیری ذراتی بزرگ که قابل بلعیده شدن توسط یک سلول واحد نیستند احساس شود، سلولهای اپیتلیویید میتوانند به یکدیگر متصل شده و تشکیل سلولهای چند هستهای به نام Gaint cell را بدهند.
اعمال ماکروفاژها
ماکروفاژها علاوه بر دربرگیری مواد بیگانه ، دارای اعمال مهم دیگری نیز در ارتباط با دفاع بدن میباشند. این سلولهای علاوه بر به انجام رساندن فاگوسیتوز ، عهدهدار ترشح فاکتورهایی هستند که باعث ایجاد تب شده و برروی پاسخهای التهابی نیز تاثیر میگذارند. آنها همچنین مسوول پردازش آنتیژن برای ایجاد پاسخ ایمنی بوده و بالاخره باعث تقویت فرآیند ترمیم بافتها میگردند. در اثر تحریک ماکروفاژ توسط باکتریها ، فرآوردههای باکتریایی و یا تخریب بافتی ، از این سلولها ، پروتیینی موسوم به اینترلوکین 1 ترشح میشود که باعث ایجاد یک پاسخ عمومی به جراحت میگردد.برخی از اعمال اینترلوکین 1 عبارتند از ایجاد تب ، تحریک نوتروفیلها و تاثیر بر روی راههای متابولیکی بدن از طریق بسیج کردن منابع انرژی به منظور از بین بردن عامل مهاجم. ماکروفاژها یکی از عناصر فعال موثر بر فرآیند التهاب هستند. این سلولها به سمت محل تهاجم میکروب جلب شده و علاوه بر کمک به حذف عامل مهاجم فاکتورهایی را نیز از خود ترشح میکنند. ماکروفاژها با داشتن شبکه آندوپلاسمی خشن ، قادر به سنتز و ترشح پروتئینها میباشند. برخی از این پروتئینها بطور مداوم آزاد میشوند. مانند آنزیم لیزوزیم و بعضی از تولیدات ماکروفاژها تنها در حین فاگوسیتوز آزاد میشوند که این ترکیبات باعث تخریب بافتی شده و تاثیر بسزایی بر التهاب دارند.
گیرندههای سطحی ماکروفاژها
ماکروفاژها دارای گیرندههای سطحی مختلفی هستند بر روی سطح ماکروفاژهای انسان و موش گیرندههایی برای آنتیبادیها وجود دارد. در نتیجه ذراتی که بوسیله آنتیبادی پوشیده شدهاند میتوانند خیلی محکم به ماکروفاژها متصل شوند و آنتیژن را در درون ماکروفاژ به این ترتیب نابود میگردد. |
پردازش آنتیژن توسط ماکروفاژها
اگر تمام مواد بیگانه بطور کامل توسط سلولهای فاگوسیتیک بالغ و هضم و نابود میشدند، دیگر نه محرکی برای ایجاد پاسخ ایمنی وجود داشت و نه نیازی به آن . بنابراین مقداری از آنتیژن باید به صورت دست نخورده برای تحریک سلولهای حساس به آنتیژن حفظ شود. آزمایشهایی که با استفاده از آنتیژن نشاندار شده توسط مواد رادیواکتیو انجام شده نشاندهنده این حقیقت است که با وجود هضم و از بین رفتن قسمت اعظم آنتیژن ، چند مولکول از آن در داخل تعدادی ماکروفاژ ، دست نخورده باقی مانده و بر روی غشای سطحی سلول یافت میشوند. همه ماکروفاژها قادر به پردازش آنتیژن برای پاسخ ایمنی نیستند.
برگرفته از :
+ نوشته شده در چهارشنبه نهم اسفند ۱۳۸۵ ساعت توسط سیامک علی حیدری
|